Di truyền học – Wikipedia tiếng Việt
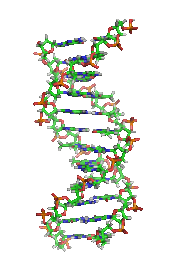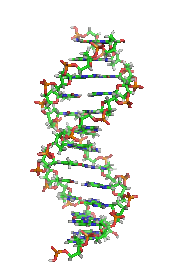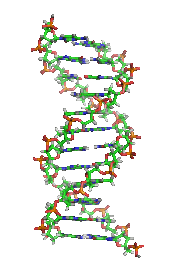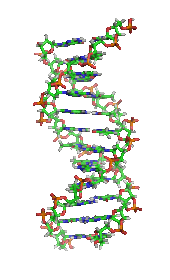 DNA, cơ sở phân tử của di truyền. Mỗi sợi DNA là một chuỗi các nucleotide, liên kết với nhau ở chính giữa có dạng như những nấc thang trong một chiếc thang xoắn.
DNA, cơ sở phân tử của di truyền. Mỗi sợi DNA là một chuỗi các nucleotide, liên kết với nhau ở chính giữa có dạng như những nấc thang trong một chiếc thang xoắn.
Di truyền học là một bộ môn sinh học, nghiên cứu về tính di truyền và biến dị ở các sinh vật[1][2].tiền sử, thực tế về việc các sinh vật sống thừa hưởng những đặc tính từ bố mẹ đã được ứng dụng để tăng sản lượng cây trồng và vật nuôi, thông qua quá trình sinh sản chọn lọc hay chọn lọc nhân tạo. Tuy nhiên, di truyền học hiện đại, tìm hiểu về quá trình di truyền, chỉ được ra đời vào khoảng cuối thế kỷ 19 với những công trình của Gregor Mendel[3]. Dù không hiểu về nền tảng vật chất của tính di truyền, Mendel vẫn nhận biết được rằng sinh vật thừa kế những tính trạng theo một cách riêng rẽ – và tiên đoán đơn vị cơ bản của quá trinh di truyền là nhân tố di truyền mà nay gọi là gen.
Mỗi gen là một đoạn xác lập của phân tử DNA, một cao phân tử sinh học được cấu thành từ bốn loại đơn phân nucleotide ; chuỗi nucleotide này mang thông tin di truyền ở sinh vật. DNA trong điều kiện kèm theo tự nhiên có dạng chuỗi xoắn kép, trong đó nucleotide ở mỗi chuỗi link bổ trợ với nhau. Mỗi chuỗi lại hoàn toàn có thể hoạt động giải trí như một khuôn để tổng hợp một chuỗi bổ trợ mới – đó là phương pháp tự nhiên tạo nên những bản sao của gen mà hoàn toàn có thể được di truyền lại cho đời sau. Chuỗi nucleotide trong gen hoàn toàn có thể được phiên mã và dịch mã trong tế bào để tạo nên chuỗi những amino acid, gọi là pôlypeptit, từ đó hình thành protein là cơ sở vật chất trực tiếp hình thành nên tính trạng ( đặc thù ) của sinh vật. Trình tự của những amino acid trong pôlypeptit của một protein tương ứng với trình tự của những nucleotide trong gen. Trình tự này được biết với tên mã di truyền. Trình tự của những nucleotide xác lập không chỉ xác lập trình tự những amino acid trong protein bậc I, mà từ đó còn xác lập cấu trúc bậc cao hơn là protein bậc II và protein bậc III và bậc IV ( nếu là đa protein ) gọi là cấu trúc ba chiều của phân tử protein 3D. Bậc cấu trúc 3D này mới giúp protein có công dụng sinh học trong tế bào sống .Protein thực thi hầu hết những công dụng thiết yếu trong mọi hoạt động giải trí sống của tế bào. Một đổi khác nhỏ của gen thường dẫn đến biến hóa trình tự amino acid, do đó dẫn đến đổi khác cấu trúc và công dụng của protein, thường gây ra đột biến hoàn toàn có thể tác động ảnh hưởng không nhỏ lên tế bào cũng như hàng loạt khung hình sống. Tuy gen đóng một vai trò to lớn trong sự hình thành tính trạng và mọi hoạt động giải trí của sinh vật, nhưng ảnh hưởng tác động của môi trường tự nhiên bên ngoài và cả những gì sinh vật đã trải qua cũng có vai trò rất quan trọng, thậm chí còn tạo ra hiệu quả sau cuối của bộc lộ tính trạng. Chẳng hạn, nhiều gen cùng lao lý chiều cao của một người, nhưng chính sách dinh dưỡng, rèn luyện của người đó cũng có ảnh hưởng tác động không nhỏ .
 [4][5].
[4][5].
Gregor Mendel, “cha đẻ của di truyền học”
Bạn đang đọc: Di truyền học – Wikipedia tiếng Việt
Khoa học di truyền được khởi đầu với công trình ứng dụng và lý thuyết của Mendel về sinh học di truyền từ giữa thế kỷ 19, tuy nhiên trước đó vẫn tồn tại những quan điểm khác nhau về di truyền. Từ thế kỷ V trước Công nguyên, Hippocrates và Aristoteles đã lần lượt đưa ra những lý thuyết của riêng mình, mà đã có ảnh hưởng không nhỏ đến các học thuyết khác sau đó. Nếu Hippocrates cho rằng các vật liệu sinh sản (“humor”) được thu thập từ tất cả các phần của cơ thể và truyền cho thế hệ con, thì Aristoteles lại phản bác lại ý kiến này, nêu ra rằng sự sinh sản bắt nguồn từ chất dinh dưỡng, trên con đường đi tới các bộ phận cơ thể thì bị chệch tới phần sinh sản, và bản chất các chất này vốn đã quy định cho cấu tạo các phần cơ thể khác nhau[6][7]. Ở thời kỳ Mendel sống, một thuyết phổ biến là quan niệm về di truyền hòa hợp (blending): cho rằng các cá thể thừa kế từ bố mẹ một hỗn hợp pha trộn các tính trạng, ví dụ như lai cây hoa đỏ với hoa trắng sẽ cho ra hoa hồng[7]. Nghiên cứu của Mendel đã bác bỏ điều này, chỉ ra tính trạng là sự kết hợp các gen độc lập với nhau hơn là một hỗn hợp liên tục. Một thuyết khác cũng nhận sự ủng hộ thời đó là sự di truyền các tính trạng tập nhiễm: tin rằng sinh vật thừa kế những tính trạng đã được biến đổi do quá trình luyện tập và nhiễm ở bố mẹ. Học thuyết này, chủ yếu gắn với Jean-Baptiste Lamarck, hiện nay không được di truyền học hiện đại thừa nhận[8]; khi sự tập nhiễm của cá thể thực tế không ảnh hưởng đến các gen mà chúng truyền cho con cái[9]. Bên cạnh đó, Charles Darwin đề ra thuyết pangen (thuyết mầm, pangenesis), có sự tương đồng với quan niệm của Hippocrates, cho rằng có các gemmule (mầm), tập trung từ các tế bào trong cơ thể về cơ quan sinh dục để thụ tinh, và con cái sinh ra chịu ảnh hưởng từ cả di truyền lẫn tính trạng tập nhiễm. Thí nghiệm của Francis Galton kiểm chứng thuyết pangen của Darwin, cho thấy rằng các gemmule ít nhất không xuất hiện trong máu thỏ[10]. Đến tận cuối thế kỷ 19, ngay cả sau khi tác phẩm của Mendel đã công bố, hiểu biết của giới khoa học về tính di truyền vẫn còn ít ỏi và chưa thực sự đúng đắn[11].
Mục Lục
Di truyền học Mendel và cổ xưa[sửa|sửa mã nguồn]
 Drosophila cho phép ông đưa ra giả thuyết rằng gen nằm trên nhiễm sắc thể.Quan sát của Morgan về sự di truyền link giới tính của đột biến mắt trắng ởcho phép ông đưa ra giả thuyết rằng gen nằm trên nhiễm sắc thể .
Drosophila cho phép ông đưa ra giả thuyết rằng gen nằm trên nhiễm sắc thể.Quan sát của Morgan về sự di truyền link giới tính của đột biến mắt trắng ởcho phép ông đưa ra giả thuyết rằng gen nằm trên nhiễm sắc thể .
Gregor Johann Mendel, một linh mục người Áo sống tại Brno (Séc; tên tiếng Đức là Brünn), đã tiến hành thí nghiệm về tính di truyền ở 7 tính trạng trên cây đậu Hà Lan từ năm 1856 đến 1863. Các nghiên cứu của ông sau đó được công bố trong bài báo “Versuche über Pflanzenhybriden” (Các thí nghiệm lai ở thực vật) tại Hội Lịch sử Tự nhiên của Brno năm 1865[12]. Cách nghiên cứu của ông là cho nhân giống theo từng tính trạng, sử dụng toán học để đánh giá số lượng và từ đó rút ra quy luật di truyền[13]. Dù các quy luật này chỉ quan sát được cho số ít tính trạng, nhưng Mendel vẫn tin rằng sự di truyền là riêng rẽ, không phải tập nhiễm, và tính di truyền của nhiều tính trạng có thể được diễn giải thông qua các quy luật và tỷ lệ đơn giản.
Tầm quan trọng của công trình Mendel không được nhận biết rộng rãi cho tới năm 1900, tức sau khi ông mất; trong năm đó, cả ba nhà khoa học Hugo de Vries (Hà Lan), Erich von Tschermak (Áo) và Carl Correns (Đức) đã nghiên cứu độc lập với nhau và cùng tái phát hiện các quy luật Mendel[14]. Năm 1900 đánh dấu một mốc khởi đầu mới cho sự phát triển của di truyền học. Năm 1905, William Bateson, một người ủng hộ Mendel, đã đặt ra thuật ngữ genetics (di truyền học).[15][16] (Tính từ genetic, xuất phát từ tiếng Hy Lạp, genesis – γένεσις, “nguồn gốc” và từ genno γεννώ, “sinh ra”, có trước danh từ này và được sử dụng lần đầu trong sinh học từ năm 1860)[17]. Bateson đã phổ biến cách dùng của từ genetics để miêu tả ngành khoa học nghiên cứu về di truyền trong bài phát biểu khai mạc Hội nghị Quốc tế lần thứ ba về lai giống cây trồng tại London, Anh năm 1906[18]. Riêng thuật ngữ gen, vốn đã được Hugo de Vries định nghĩa với tên gọi pangen từ năm 1889 là: “phần tử nhỏ nhất [đại diện cho] một đặc điểm di truyền”[19], được Wilhelm Johannsen giới thiệu lại trong các tác phẩm của ông vào hai thập niên sau đó – trong đó ông cũng nêu ra thuật ngữ kiểu gen (genotype) và kiểu hình (phenotype)[20][21].
Sau sự tái phát hiện khu công trình của Mendel, những nhà khoa học đã cố gắng nỗ lực xác lập những phân tử nào trong tế bào tiếp đón tính di truyền. Trước đó, nhiễm sắc thể đã được phát hiện, và những quan điểm tiên phong về di truyền nhiễm sắc thể đã được đưa ra [ 22 ], phải kể đến là thuyết di truyền nhiễm sắc thể của August Weismann năm 1892 [ 23 ] và giả thuyết gắn những tác nhân Mendel với nhiễm sắc thể của Walter Sutton năm 1903 [ 24 ]. Năm 1910, Thomas Hunt Morgan chứng minh và khẳng định rằng gen nằm trên nhiễm sắc thể, dựa trên sự quan sát về đột biến mắt trắng ở ruồi giấm [ 25 ]. Năm 1913, một sinh viên của ông, Alfred Sturtevant đã sử dụng hiện tượng kỳ lạ di truyền link để chỉ ra rằng gen được sắp xếp theo đường thẳng ( tuyến tính ) trên nhiễm sắc thể, và kiến thiết xây dựng nên map link gen tiên phong [ 26 ] .
Di truyền học phân tử[sửa|sửa mã nguồn]
Dù sự sống sót của gen trên nhiễm sắc thể – hợp thành từ protein và DNA – đã được xác nhận, tuy nhiên người ta vẫn chưa biết đến cái gì trong hai chất đó đóng vai trò di truyền. Năm 1928, Frederick Griffith mày mò ra hiện tượng kỳ lạ biến nạp : những vi trùng đã chết hoàn toàn có thể chuyển vật tư di truyền của chúng để làm đổi khác những vi trùng còn sống khác [ 27 ]. Năm 1944, Oswald Theodore Avery, Colin McLeod và Maclyn McCarty đã thực thi thí nghiệm trực tiếp xác lập DNA là phân tử tiếp đón biến nạp [ 28 ]. Tuy nhiên, đến tận năm 1952, thí nghiệm Hershey – Chase mới cho thấy DNA ( chứ không phải protein ) là vật tư di truyền của virus xâm nhiễm vi trùng, cung ứng thêm dẫn chứng chứng tỏ DNA là phân tử tiếp đón tính năng di truyền [ 29 ] .James D. Watson và Francis Crick cho sinh ra quy mô cấu trúc DNA năm 1953, sử dụng khu công trình tinh thể học tia X của Rosalind Franklin, chứng tỏ rằng DNA có cấu trúc xoắn kép [ 30 ] [ 31 ]. Mô hình DNA của họ gồm có hai chuỗi với những nucleotide phía trong, mỗi một nucleotide link bổ trợ với một nucleotide ở chuỗi khác tạo thành hình dạng giống như thanh ngang trên một chiếc thang xoắn [ 32 ]. Cấu trúc này chỉ ra rằng thông tin di truyền sống sót trên dãy nucleotide ở mỗi chuỗi DNA, và cũng đưa ra gợi ý về một phương pháp nhân đôi đơn thuần : nếu chuỗi kép bị tách rời, chuỗi bổ trợ mới hoàn toàn có thể được tái dựng lại từ mỗi chuỗi đơn cũ .
Dù cấu trúc DNA cho thấy được cách thức di truyền, người ta vẫn chưa biết rõ ràng về cách mà DNA ảnh hưởng lên hoạt động của tế bào. Trước đấy, năm 1941, George Wells Beadle và Edward Lawrie Tatum đã đề ra thuyết “một gen-một enzym”, chứng minh vai trò điều khiển và điều hòa của gen lên các phản ứng sinh hóa ở mốc bánh mỳ Neurospora[33][34], đồng thời phương pháp của họ – ứng dụng di truyền học vào sinh hóa ở vi sinh vật – cũng mở ra một phạm vi nghiên cứu mới ngay sau đó[35]. Trong những năm sau đó, các nhà khoa học đã cố gắng tìm ra cách DNA điều khiển quá trình tổng hợp protein. Họ đã khám phá được rằng tế bào đã sử dụng DNA như một khuôn để tạo nên phân tử RNA thông tin tương ứng. Dãy nucleotide trên RNA thông tin lại tiếp tục được sử dụng để tạo nên dãy amino acid ở protein; trình tự của dãy nucleotide được dịch mã để tạo thành dãy amino acid được gọi là mã di truyền. Nó được dựa trên sự sắp xếp những bộ ba base nitơ không chồng lấn nhau, gọi là codon, mỗi codon mã hóa cho một amino acid. Điều này lần đầu tiên được miêu tả trong thí nghiệm của Crick, Brenner và các cộng sự năm 1961[36]. Trong những năm 1961-1966 đã ghi nhận kết quả nỗ lực của các nhà khoa học để giải mã được toàn bộ 64 codon, chủ yếu là những công trình do nhóm của M. Nirenberg (thí nghiệm Nirenberg và Matthaei) và nhóm của H. Khorana thực hiện.
Những hiểu biết mới tầm phân tử về tính di truyền đã tạo nên sự bùng nổ trong nghiên cứu và điều tra. Một bước tăng trưởng quan trọng là chiêu thức xác lập trình tự DNA gián đoạn chuỗi năm 1977 của Frederick Sanger : công nghệ tiên tiến này được cho phép những nhà khoa học đọc được trình tự nucleotide trên một phân tử DNA [ 37 ]. Năm 1983, Kary Banks Mullis tăng trưởng phản ứng chuỗi trùng hợp ( PCR ), cung ứng một chiêu thức nhanh gọn để phân lập và khuếch đại một đoạn DNA riêng không liên quan gì đến nhau từ một hỗn hợp [ 38 ]. Những cố gắng nỗ lực chung trong Dự án Bản đồ gen Người và nỗ lực song song của công ty tư nhân Celera Genomics, cũng như những công nghệ tiên tiến khác, ở đầu cuối đã thành công xuất sắc trong việc xác lập trình tự bộ gen người vào năm 2003 [ 39 ] .
Đặc trưng của di truyền[sửa|sửa mã nguồn]
Di truyền riêng rẽ và quy luật Mendel[sửa|sửa mã nguồn]

Ký hiệu và biểu đồ[sửa|sửa mã nguồn]
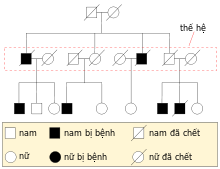 Một sơ đồ phả hệ giúp theo dõi được kiểu di truyền của một tính trạng đã cho .Các nhà di truyền học sử dụng những biểu đồ và hình tượng để miêu tả sự di truyền. Một gen được biểu trưng bởi một ( hay vài ) ký tự — trong đó ký tự viết hoa tượng trưng cho allele trội và ký tự viết thường tượng trưng cho allele lặn [ 43 ]. Thông thường hình tượng ” + ” được sử dụng để bộc lộ allele thường, không đột biến ở một gen .Ở những thí nghiệm lai và thụ tinh ( đặc biệt quan trọng về những quy luật Mendel ), cha mẹ được xem là thế hệ ” P “, con cháu của chúng được gọi là thế hệ ” F1 ” ( ” first filial ” ). Khi những thành viên F1 giao phối với nhau, con của F1 lại liên tục gọi là ” F2 “. Một trong những biểu đồ thường được sử dụng để Dự kiến hiệu quả lai là bảng Punnett ( do Reginald Punnett phát minh sáng tạo ) .Khi nghiên cứu và điều tra về những bệnh di truyền ở người, những nhà di truyền học thường dùng sơ đồ phả hệ để diễn đạt sự di truyền ở những tính trạng [ 44 ]. Các sơ đồ này sẽ sắp xếp sự di truyền của một tính trạng trên một cây phả hệ .
Một sơ đồ phả hệ giúp theo dõi được kiểu di truyền của một tính trạng đã cho .Các nhà di truyền học sử dụng những biểu đồ và hình tượng để miêu tả sự di truyền. Một gen được biểu trưng bởi một ( hay vài ) ký tự — trong đó ký tự viết hoa tượng trưng cho allele trội và ký tự viết thường tượng trưng cho allele lặn [ 43 ]. Thông thường hình tượng ” + ” được sử dụng để bộc lộ allele thường, không đột biến ở một gen .Ở những thí nghiệm lai và thụ tinh ( đặc biệt quan trọng về những quy luật Mendel ), cha mẹ được xem là thế hệ ” P “, con cháu của chúng được gọi là thế hệ ” F1 ” ( ” first filial ” ). Khi những thành viên F1 giao phối với nhau, con của F1 lại liên tục gọi là ” F2 “. Một trong những biểu đồ thường được sử dụng để Dự kiến hiệu quả lai là bảng Punnett ( do Reginald Punnett phát minh sáng tạo ) .Khi nghiên cứu và điều tra về những bệnh di truyền ở người, những nhà di truyền học thường dùng sơ đồ phả hệ để diễn đạt sự di truyền ở những tính trạng [ 44 ]. Các sơ đồ này sẽ sắp xếp sự di truyền của một tính trạng trên một cây phả hệ .
Tương tác của nhiều gen[sửa|sửa mã nguồn]
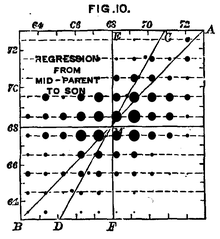 Chiều cao con người là một tính trạng di truyền phức tạp. Các tài liệu của Francis Galton từ 1889 cho thấy : sự liên hệ toàn bộ chiều cao của con cháu là một hàm trung bình của chiều cao cha mẹ. Trong khi mức biến hóa đối sánh tương quan, còn lại trong chiều cao con cháu bộc lộ rằng môi trường tự nhiên cũng là một tác nhân quan trọng ở tính trạng này .Mỗi sinh vật có hàng ngàn gen và ở những sinh vật sinh sản hữu tính, sự phân ly những gen này nhìn chung độc lập với nhau. Điều này có nghĩa là sự di truyền của một allele tính trạng hạt đậu vàng hay xanh không có tương quan tới sự di truyền của cặp allele màu hoa trắng hoặc tía. Hiện tượng này, được biết đến là Quy luật thứ hai của Mendel hay ” Quy luật phân ly độc lập “, mang ý nghĩa : những allele của những gen khác nhau sẽ đổi khác ngẫu nhiên khi phân ly từ cha mẹ và sẽ tạo ra thế hệ con với nhiều tổng hợp gen khác nhau. Dù thế, 1 số ít gen lại không phân ly độc lập với nhau, biểu lộ tính link gen .
Chiều cao con người là một tính trạng di truyền phức tạp. Các tài liệu của Francis Galton từ 1889 cho thấy : sự liên hệ toàn bộ chiều cao của con cháu là một hàm trung bình của chiều cao cha mẹ. Trong khi mức biến hóa đối sánh tương quan, còn lại trong chiều cao con cháu bộc lộ rằng môi trường tự nhiên cũng là một tác nhân quan trọng ở tính trạng này .Mỗi sinh vật có hàng ngàn gen và ở những sinh vật sinh sản hữu tính, sự phân ly những gen này nhìn chung độc lập với nhau. Điều này có nghĩa là sự di truyền của một allele tính trạng hạt đậu vàng hay xanh không có tương quan tới sự di truyền của cặp allele màu hoa trắng hoặc tía. Hiện tượng này, được biết đến là Quy luật thứ hai của Mendel hay ” Quy luật phân ly độc lập “, mang ý nghĩa : những allele của những gen khác nhau sẽ đổi khác ngẫu nhiên khi phân ly từ cha mẹ và sẽ tạo ra thế hệ con với nhiều tổng hợp gen khác nhau. Dù thế, 1 số ít gen lại không phân ly độc lập với nhau, biểu lộ tính link gen .
Thực tế, các gen khác nhau lại có thể tương tác với nhau theo một cách nào đấy và ảnh hưởng lên một tính trạng chung. Một ví dụ là ở loài hoa Omphalodes verna, tồn tại một gen với hai allele xác định tính trạng màu hoa: xanh lam hoặc đỏ tía. Một gen khác điều khiển khả năng có màu của hoa: có màu hoặc không màu (màu trắng). Khi một cây có hai allele hoa trắng, hoa của cây đấy luôn màu trắng – cho dù gen đầu tiên có allele hoa đỏ hay xanh. Sự tương tác được gọi là tương tác át chế (epistasis), khi gen thứ hai át chế sự biểu hiện của gen thứ nhất[45]. Bên cạnh tương tác át chế còn có kiểu tương tác bổ trợ, nghĩa là sự biểu hiện của hai hay nhiều allele ở các gen khác nhau sẽ tạo kiểu hình mới, khác với kiểu hình riêng được biểu hiện khi có mặt các allele ở từng gen riêng lẻ[46].
Có nhiều tính trạng không riêng rẽ (ví dụ hoa màu trắng/tía) mà thay vì thế lại biểu hiện liên tục (ví dụ chiều cao và màu da). Các tính trạng phức hợp này được tạo bởi tác động cộng gộp của nhiều gen[47]. Sự chi phối của các gen này là tương đương, có vai trò biến đổi mức độ biểu hiện, bên cạnh điều kiện môi trường của sinh vật. Tỷ lệ mà các gen của sinh vật đóng góp cho một tính trạng phức hợp được gọi là mức di truyền (heritability)[48]. Số đo của mức di truyền chỉ là tương đối – khi môi trường càng dễ biến đổi sẽ càng tác động lớn hơn lên toàn bộ mức thay đổi của tính trạng. Một ví dụ, chiều cao của con người là một tính trạng phức hợp với mức di truyền là 89% tại Mỹ. Ở Nigeria, nơi người dân có điều kiện dinh dưỡng và y tế hay thay đổi hơn, chiều cao của họ có mức di truyền chỉ là 62%[49].
Cơ sở phân tử của tính di truyền[sửa|sửa mã nguồn]
DNA và nhiễm sắc thể[sửa|sửa mã nguồn]
 Biểu đồ năm 1882 của Walther Flemming về sự phân loại tế bào nhân chuẩn. Các nhiễm sắc thể được nhân đôi, co xoắn và tổ chức triển khai lại. Sau đó, khi tế bào phân loại, những nhiễm sắc thể tương đương cũng phân làn vào những tế bào con .
Biểu đồ năm 1882 của Walther Flemming về sự phân loại tế bào nhân chuẩn. Các nhiễm sắc thể được nhân đôi, co xoắn và tổ chức triển khai lại. Sau đó, khi tế bào phân loại, những nhiễm sắc thể tương đương cũng phân làn vào những tế bào con .
- Các vi khuẩn (nhân sơ) thường là đơn bội (mức bội thể bằng 1) chỉ có duy nhất một mỗi nhiễm sắc thể. Còn tuyệt đại các sinh vật khác có bộ nhiễm sắc thể lưỡng bội (mức bội thể bằng 2), trong đó mỗi nhiễm sắc thể có một nhiễm sắc thể khác giống nó về hình dạng, kích thước và trình tự lô-cut gen giống nó, tạo thành cặp nhiễm sắc thể tương đồng, do đó có hai bản sao cho mỗi gen: 1 nhận từ bố, còn 1 nhận từ mẹ [55]. Các alen (allele) của cùng một gen nằm trên vị trí như nhau gọi là locus.
- Ở các loài có nhiễm sắc thể giới tính, thì cặp nhiễm sắc thể giới tính (kí hiệu là X và Y, hoặc W và Z) giữ vai trò xác định giới tính của mỗi cá thể[56]. Đối với cặp nhiễm sắc thể giới tính này, thì lúc tương đồng (như X với X), lúc không tương đồng (như X với Y), do nhiễm sắc thể Y/Z rất ngắn và có rất ít gen. Sự biến đổi bất thường về số lượng nhiễm sắc thể giới tính luôn dẫn tới những rối loạn về giới tính và về di truyền, gây bệnh khác thường.
Khi tế bào phân chia, toàn thể bộ gen đều được sao chép và mỗi tế bào con đều nhận được một bản sao bộ gen của tế bào mẹ. Quá trình này gọi là nguyên phân, là dạng đơn giản nhất của sinh sản và là cơ sở của sự sinh sản vô tính. Sinh sản vô tính có thể xuất hiện ở cả các sinh vật đa bào, tạo ra thế hệ con thừa hưởng bộ gen từ chỉ duy nhất một cá thể mẹ. Thế hệ con mà nhìn chung giống hệt như mẹ được gọi là dòng vô tính (clone).
Sinh vật nhân chuẩn thường sinh sản hữu tính để tạo ra con cháu có vật tư di truyền lấy từ hai bộ gen khác nhau của cả hai thành viên cha mẹ. Quá trình sinh sản hữu tính luân phiên nhau giữa hai dạng đơn bội ( 1 n ) và lưỡng bội ( 2 n ) [ 56 ]. Các tế bào đơn bội phối hợp vật chất di truyền của chúng, tạo ra tế bào lưỡng bội với những nhiễm sắc thể ghép cặp. Các sinh vật lưỡng bội tạo ra thể đơn bội bằng cách phân loại, tạo ra tế bào con thừa kế ngẫu nhiên một nhiễm sắc thể trong mỗi cặp nhiễm sắc thể tương đương. Đa phần động vật hoang dã và nhiều thực vật ở thể lưỡng bội trong hầu hết vòng đời, với thể đơn bội đổi khác thành những giao tử đơn bào. Một quy trình đa phần trong sinh sản hữu tính là giảm phân, quy trình phân bào chuyên biệt diễn ra ở tế bào sinh dục, trong đó bộ nhiễm sắc thể lưỡng bội được nhân đôi, trải qua hai lần phân bào để tạo nên những tế bào con đơn bội có số lượng nhiễm sắc thể giảm đi 50% so với tế bào mẹ bắt đầu [ 57 ] .Dù không sử dụng chính sách sinh sản hữu tính kiểu lưỡng bội / đơn bội như trên, vi trùng vẫn có nhiều cách để thu nhận thông tin di truyền. Một số vi trùng có năng lực tiếp hợp, chuyển một vòng nhỏ DNA tới một vi trùng khác [ 58 ]. Vi khuẩn cũng hoàn toàn có thể lấy những đoạn DNA thô từ môi trường tự nhiên và tích hợp chúng vào trong bộ gen của vi trùng, hiện tượng kỳ lạ này được biết đến là sự biến nạp [ 59 ]. Quá trình này hoàn toàn có thể dẫn đến sự chuyển gen ngang, truyền những đoạn thông tin di truyền giữa những sinh vật không có mối liên hệ với nhau .
 Minh họa năm 1916 của Thomas Hunt Morgan về quy trình trao đổi chéo kép giữa hai nhiễm sắc thể tương đương .
Minh họa năm 1916 của Thomas Hunt Morgan về quy trình trao đổi chéo kép giữa hai nhiễm sắc thể tương đương .
Tái tổng hợp và link gen[sửa|sửa mã nguồn]
Trạng thái lưỡng bội tự nhiên của bộ nhiễm sắc thể được cho phép gen nằm trên những nhiễm sắc thể khác nhau phân ly độc lập với nhau trong quy trình sinh sản hữu tính, tái tổng hợp tạo tổng hợp gen mới. Gen trên cùng một nhiễm sắc thể về triết lý không khi nào tái tổng hợp [ 60 ], tuy nhiên, thực tiễn vẫn diễn ra do Open quy trình trao đổi chéo nhiễm sắc thể. Khi trao đổi chéo, hai nhiễm sắc thể trao đổi những đoạn DNA cho nhau, và đổi chỗ những allele giữa hai nhiễm sắc thể [ 61 ]. Trao đổi chéo thường thì diễn ra vào kỳ trước I ( kỳ đầu của lần phân loại đầu ) của quy trình giảm phân .Xác suất trao đổi chéo giữa hai điểm đã cho trên nhiễm sắc thể có tương quan đến khoảng cách giữa chúng. Ở một khoảng cách dài tùy ý, Tỷ Lệ trao đổi chéo đủ cao để sự di truyền những gen diễn ra tương đối riêng rẽ. Tuy nhiên, với những gen gần nhau hơn, Tỷ Lệ trao đổi chéo thấp chứng tỏ những gen có tính link di truyền – allele của hai gen này có khuynh hướng di truyền gắn liền với nhau. Các chỉ số về tính link của chuỗi nhiều gen hoàn toàn có thể được phối hợp tạo nên một map link, giúp xác lập gần đúng vị trí sắp xếp những gen trên nhiễm sắc thể [ 62 ] .
Biểu hiện gen[sửa|sửa mã nguồn]
Mã di truyền[sửa|sửa mã nguồn]
 Mã di truyền : DNA, qua một trung gian RNA thông tin, mã hóa cho protein với những bộ ba mã hóa .Gen nhìn chung biểu lộ ảnh hưởng tác động của chúng trải qua việc tổng hợp protein, những phân tử phức tạp đảm nhiệm hầu hết tính năng trong tế bào. Protein là một chuỗi những amino acid ; trình tự DNA của một gen, trải qua trung gian RNA thông tin ( mRNA ), được sử dụng để tạo nên trình tự phân tử protein riêng không liên quan gì đến nhau. Quá trình này khởi đầu với việc tổng hợp một phân tử mRNA với trình tự tương ứng trình tự DNA của gen quá trình này gọi là phiên mã .Phân tử mRNA sau đó lại được sử dụng như một khuôn để tạo thành trình tự amino acid tương ứng trải qua một quy trình gọi là dịch mã. Mỗi bộ ba nucleotide ( codon ) ở dãy này tương ứng với một trong 20 loại amino acid xuất hiện trong protein – sự tương ứng này gọi là mã di truyền [ 63 ]. Dòng thông tin đi theo một hướng duy nhất : thông tin khi được truyền từ chuỗi nucleotide tới chuỗi amino acid của protein, nó không khi nào được truyền ngược lại từ protein tới chuỗi DNA – hiện tượng kỳ lạ này được Francis Crick gọi là ” luận thuyết TT của sinh học phân tử ” [ 64 ] .Trình tự đặc hiệu của amino acid dẫn đến cấu trúc ba chiều độc nhất của protein, và điều này lại tương quan đến công dụng của protein [ 65 ] [ 66 ]. Một số protein là những phân tử có cấu trúc đơn thuần, ví dụ như collagen tạo nên những sợi mô. Protein cũng hoàn toàn có thể kết nối với nhau và với những phân tử đơn thuần khác, nhiều khi hoạt động giải trí như những enzym xúc tác cho phản ứng hóa học của những phân tử kết nối với nó ( những phản ứng không làm đổi khác cấu trúc của bản thân protein ). Cấu trúc của protein có tính động, ví dụ hemoglobin hoàn toàn có thể quy đổi thành những dạng độc lạ đôi chút mỗi khi protein này thu nhận, luân chuyển và giải phóng oxy trong máu động vật hoang dã có vú .Chỉ một biến hóa nucleotide trong DNA cũng hoàn toàn có thể gây biến hóa trình tự amino acid trong protein. Bởi cấu trúc protein là hiệu quả của trình tự amino acid, nên những đổi khác trình tự hoàn toàn có thể làm biến hóa bất ngờ đột ngột những đặc tính của protein, do sự mất không thay đổi cấu trúc hay đổi khác mặt phẳng protein là nguyên do dẫn tới đổi khác về tính tương tác của nó với những protein và phân tử khác. Một ví dụ, bệnh thiếu máu hồng cầu liềm là một bệnh di truyền ở người, gây ra do độc lạ một base trong vùng mã hóa phần β-globin của hemoglobin, khiến một amino acid ở protein này cũng đổi khác theo và làm biến hóa đặc tính vật lý của hemoglobin [ 67 ]. Những hemoglobin này phối hợp với nhau, làm biến hóa hình dạng tế bào hồng cầu ; những tế bào hồng cầu hình liềm không còn chuyển dời thuận tiện trong mạch máu, chúng có xu thế ùn tắc và thoái hóa, gây nên những yếu tố sức khỏe thể chất gắn liền với bệnh này .
Mã di truyền : DNA, qua một trung gian RNA thông tin, mã hóa cho protein với những bộ ba mã hóa .Gen nhìn chung biểu lộ ảnh hưởng tác động của chúng trải qua việc tổng hợp protein, những phân tử phức tạp đảm nhiệm hầu hết tính năng trong tế bào. Protein là một chuỗi những amino acid ; trình tự DNA của một gen, trải qua trung gian RNA thông tin ( mRNA ), được sử dụng để tạo nên trình tự phân tử protein riêng không liên quan gì đến nhau. Quá trình này khởi đầu với việc tổng hợp một phân tử mRNA với trình tự tương ứng trình tự DNA của gen quá trình này gọi là phiên mã .Phân tử mRNA sau đó lại được sử dụng như một khuôn để tạo thành trình tự amino acid tương ứng trải qua một quy trình gọi là dịch mã. Mỗi bộ ba nucleotide ( codon ) ở dãy này tương ứng với một trong 20 loại amino acid xuất hiện trong protein – sự tương ứng này gọi là mã di truyền [ 63 ]. Dòng thông tin đi theo một hướng duy nhất : thông tin khi được truyền từ chuỗi nucleotide tới chuỗi amino acid của protein, nó không khi nào được truyền ngược lại từ protein tới chuỗi DNA – hiện tượng kỳ lạ này được Francis Crick gọi là ” luận thuyết TT của sinh học phân tử ” [ 64 ] .Trình tự đặc hiệu của amino acid dẫn đến cấu trúc ba chiều độc nhất của protein, và điều này lại tương quan đến công dụng của protein [ 65 ] [ 66 ]. Một số protein là những phân tử có cấu trúc đơn thuần, ví dụ như collagen tạo nên những sợi mô. Protein cũng hoàn toàn có thể kết nối với nhau và với những phân tử đơn thuần khác, nhiều khi hoạt động giải trí như những enzym xúc tác cho phản ứng hóa học của những phân tử kết nối với nó ( những phản ứng không làm đổi khác cấu trúc của bản thân protein ). Cấu trúc của protein có tính động, ví dụ hemoglobin hoàn toàn có thể quy đổi thành những dạng độc lạ đôi chút mỗi khi protein này thu nhận, luân chuyển và giải phóng oxy trong máu động vật hoang dã có vú .Chỉ một biến hóa nucleotide trong DNA cũng hoàn toàn có thể gây biến hóa trình tự amino acid trong protein. Bởi cấu trúc protein là hiệu quả của trình tự amino acid, nên những đổi khác trình tự hoàn toàn có thể làm biến hóa bất ngờ đột ngột những đặc tính của protein, do sự mất không thay đổi cấu trúc hay đổi khác mặt phẳng protein là nguyên do dẫn tới đổi khác về tính tương tác của nó với những protein và phân tử khác. Một ví dụ, bệnh thiếu máu hồng cầu liềm là một bệnh di truyền ở người, gây ra do độc lạ một base trong vùng mã hóa phần β-globin của hemoglobin, khiến một amino acid ở protein này cũng đổi khác theo và làm biến hóa đặc tính vật lý của hemoglobin [ 67 ]. Những hemoglobin này phối hợp với nhau, làm biến hóa hình dạng tế bào hồng cầu ; những tế bào hồng cầu hình liềm không còn chuyển dời thuận tiện trong mạch máu, chúng có xu thế ùn tắc và thoái hóa, gây nên những yếu tố sức khỏe thể chất gắn liền với bệnh này .
Một vài gen được phiên mã tạo RNA, nhưng RNA lại không tiếp tục dịch mã thành sản phẩm protein – được gọi chung là RNA không mã hóa (non-coding RNA). Trong một số trường hợp, RNA không mã hóa lại gập uốn hình thành những cấu trúc, tham gia các chức năng then chốt của tế bào (ví dụ RNA ribosome và RNA vận chuyển). RNA cũng có thể có tác động điều hòa thông qua tương tác lai với những phân tử RNA khác (ví dụ microRNA).
Kiểu gen, kiểu hình và thiên nhiên và môi trường[sửa|sửa mã nguồn]
 Một con mèo Xiêm mang đột biến mẫn cảm nhiệt độ về tổng hợp sắc tố .
Một con mèo Xiêm mang đột biến mẫn cảm nhiệt độ về tổng hợp sắc tố .
Dù các gen chứa đựng mọi thông tin một sinh vật cần để thực hiện chức năng, môi trường vẫn đóng vai trò quan trọng trong việc xác định kiểu hình sau cùng— tính lưỡng phân trên được nói đến trong cụm từ “bản chất đối chọi môi trường” (nature vs. nurture). Kiểu hình của các sinh vật phụ thuộc vào sự tương tác giữa kiểu gen và môi trường. Một ví dụ cho kết luận này là trường hợp đột biến mẫn cảm với nhiệt độ. Thông thường, một amino acid đơn lẻ thay đổi trong chuỗi protein không làm thay đổi hoạt động và tương tác của nó với các phân tử khác, tuy nhiên điều này lại làm mất ổn định cấu trúc. Trong môi trường nhiệt độ cao, các phân tử chuyển động nhanh hơn và va chạm vào nhau, kết quả protein không còn giữ được cấu trúc và mất đi chức năng. Ở môi trường nhiệt độ thấp, cấu trúc protein lại ổn định và thực hiện chức năng bình thường. Loại đột biến này có thể quan sát thấy ở màu lông những con mèo Xiêm, khi một đột biến xảy ra ở enzym phụ trách sản xuất sắc tố, khiến enzym mất ổn định và mất chức năng ở nhiệt độ cao[68]. Protein này sẽ duy trì chức năng ở những vùng da lạnh hơn – như chân, tai, đuôi và mặt – làm cho giống mèo này vẫn có phần lông màu đen ở những vùng nói trên.
Xem thêm: ‘vậy hả’ là gì?, Từ điển Tiếng Việt
Một ví dụ khác là ảnh hưởng tác động thâm thúy của thiên nhiên và môi trường lên bệnh di truyền phenylketon niệu ở người [ 69 ]. Đột biến tạo nên chứng bệnh này, phá hoại năng lực phân giải amino acid phenylalanine, tích tụ những chất trung gian gây độc, tiếp đó gây nên những ảnh hưởng tác động rất xấu lên thần kinh. Nếu một người bị mắc đột biến phenylketon niệu đi theo một chính sách siêu thị nhà hàng khắt khe tránh xa loại amino acid này, anh ta vẫn duy trì được thực trạng thông thường và khỏe mạnh .
Điều hòa gen[sửa|sửa mã nguồn]
 Nhân tố phiên mã kết nối với DNA, ảnh hưởng tác động lên sự phiên mã của những gen đã link .
Nhân tố phiên mã kết nối với DNA, ảnh hưởng tác động lên sự phiên mã của những gen đã link .
Bộ gen của một sinh vật bao gồm hàng nghìn gen, nhưng không phải bất cứ gen nào cũng cần được hoạt động tại mọi thời điểm. Một gen chỉ có thể được biểu hiện khi nó được phiên mã thành mRNA (và dịch mã thành protein); thực tế tồn tại nhiều cách thức trong tế bào để điều khiển sự biểu hiện của gen, đảm bảo cho protein nào được sản xuất chỉ khi tế bào cần. Các nhân tố phiên mã là những protein điều hòa được gắn vào điểm khởi đầu của gen, có vai trò hoạt hóa hay ức chế sự phiên mã của gen đó[70]. Ví dụ, trong bộ gen của vi khuẩn E. coli có một dãy nhiều gen cần thiết cho việc tổng hợp amino acid tryptophan. Tuy nhiên, khi tryptophan đã sẵn có trong tế bào, những gen tổng hợp trytophan sẽ không được duy trì hoạt động. Sự có mặt của trytophan trực tiếp ảnh hưởng đến hoạt động của những gen này – những phân tử trytophan liên kết với chất ức chế trytophan (trp repressor – một nhân tố phiên mã), thay đổi cấu trúc của phân tử này giúp nó gắn được vào gen. Trytophan repressor ngăn chặn quá trình phiên mã và sự biểu hiện của các gen tổng hợp trytophan, do đó tạo nên sự điều hòa liên hệ ngược âm tính của quá trình tổng hợp loại amino acid này[71].
Những độc lạ trong bộc lộ gen đặc biệt quan trọng rõ ràng ở những sinh vật đa bào, khi những tế bào cùng có chung bộ gen nhưng lại có cấu trúc và hoạt động giải trí rất khác nhau, dựa trên sự biểu lộ của những tập hợp gen khác nhau. Tất cả tế bào trong một khung hình đa bào đều có nguồn gốc từ một tế bào duy nhất, được biệt hóa thành những dạng tế bào khác nhau khi phản ứng lại những tín hiệu ngoại và gian bào, và từ từ kiến lập những phương pháp bộc lộ gen khác nhau để triển khai những hoạt động giải trí khác nhau. Bởi không có một gen riêng không liên quan gì đến nhau nào chịu nghĩa vụ và trách nhiệm cho sự tăng trưởng những cấu trúc bên trong sinh vật đa bào, nên những phương pháp biển hiện trên đều phát sinh từ những tương tác phức tạp giữa nhiều tế bào .
Ở sinh vật nhân chuẩn, tồn tại những đặc tính cấu trúc của chromatin có ảnh hưởng đến sự phiên mã của gen, thường ở dạng thường biến (modification) trên DNA hay chromatin mà vẫn được di truyền ổn định sang các tế bào con[72]. Những đặc tính này được gọi là “ngoại di truyền” (epigenetic) bởi chúng xuất hiện ở ngoài phạm vi trình tự DNA và vẫn được duy trì từ tế bào này sang thế hệ kế tiếp. Bởi có những đặc tính ngoại di truyền, các dạng tế bào khác nhau sinh trưởng trong cùng một môi trường có thể giữ lại những đặc điểm riêng biệt của chúng. Dù các đặc tính ngoại di truyền nhìn chung mang tính động trong tiến trình phát triển và không được giữ lại ở thế hệ sau của thế hệ kế tiếp, nhưng một số, như hiện tượng cận đột biến (paramutation), vẫn được di truyền qua nhiều thế hệ và tồn tại như những ngoại lệ hiếm hoi nằm ngoài quy luật chung của DNA (được xem như cơ sở căn bản của tính di truyền)[73].
Biến đổi di truyền[sửa|sửa mã nguồn]
 Sự lặp gen ( lặp đoạn nhiễm sắc thể ) được cho phép đa dạng hóa bằng cách phân phối thêm nguyên vật liệu di truyền : một gen hoàn toàn có thể đột biến và mất đi tính năng bắt đầu mà không làm tổn hại đến sinh vật .
Sự lặp gen ( lặp đoạn nhiễm sắc thể ) được cho phép đa dạng hóa bằng cách phân phối thêm nguyên vật liệu di truyền : một gen hoàn toàn có thể đột biến và mất đi tính năng bắt đầu mà không làm tổn hại đến sinh vật .
Trong quá trình tự nhân đôi DNA, những sai sót đôi lúc diễn ra khi tổng hợp chuỗi thứ hai. Những lỗi này, gọi là đột biến, có thể có tác động lên kiểu hình của cá thể, đặc biệt nếu chúng xảy ra tại phần mã hóa protein của một gen. Tỷ lệ sai sót thường rất thấp – 1 lỗi trong 10-100 triệu base – nhờ khả năng “đọc sửa” của DNA polymerase[74][75] (Nếu không được đọc sửa, tỷ lệ lỗi sẽ cao hơn hàng nghìn lần, bởi nhiều virus dựa vào DNA hay RNA polymerase thiếu khả năng đọc sửa, làm tăng tỷ lệ đột biến lên cao). Quá trình làm tăng tỷ lệ biến đổi ở DNA được gọi là “gây đột biến” (mutagenic): các hóa chất gây đột biến đẩy mạnh làm tăng sai sót trong tái bản DNA, gây nhiễu loạn kết cấu của sự ghép cặp base; trong khi tia UV tạo ra đột biến bằng cách gây tổn hại cấu trúc DNA[76]. Các tổn thương về hóa học ở DNA cũng có thể diễn ra một cách tự nhiên, và tế bào sử dụng cơ chế sửa chữa DNA để sửa lại các ghép đôi không cân xứng hay đứt gãy ở DNA – tuy nhiên việc sửa chữa này thỉnh thoảng vẫn thất bại và không thể đưa DNA trở lại chuỗi ban đầu.
Với những sinh vật sử dụng trao đổi chéo nhiễm sắc thể để trao đổi DNA và tái tổng hợp gen, những sai sót khi bắt cặp thẳng hàng ở giảm phân cũng hoàn toàn có thể tạo ra đột biến [ 77 ]. Lỗi trong trao đổi chéo đặc biệt quan trọng xảy ra khi những phần giống nhau trên những nhiễm sắc thể khiến chúng bắt cặp nhầm lẫn, làm một số ít vùng của bộ gen bị đột biến. Những lỗi này tạo nên sự biến hóa cấu trúc lớn trong nhiễm sắc thể và trình tự DNA – dẫn đến sự lặp đoạn, hòn đảo đoạn hay mất đoạn của tổng thể những vùng trên, hoặc sự hoán đổi ngẫu nhiên những đoạn giữa những nhiễm sắc thể khác nhau ( được gọi là chuyển đoạn ) .
Chọn lọc tự nhiên và tiến hóa[sửa|sửa mã nguồn]
Đột biến tạo nên các cá thể với kiểu gen khác nhau, và những khác biệt này dẫn tới những kiểu hình khác nhau. Nhiều đột biến có tác động không lớn lên kiểu hình, sức khỏe và sự thích ứng sinh sản của sinh vật. Tác động của đột biến thường là có hại, nhưng đôi khi lại trở nên có ích. Những nghiên cứu trên ruồi giấm Drosophila melanogaster cho thấy nếu một đột biến thay đổi một protein mã hóa bởi một gen, điều này hầu như sẽ gây tác hại: 70% trong những đột biến này là có hại, số còn lại là trung tính hoặc có lợi nhưng rất thấp[78].
Di truyền học quần thể nghiên cứu về sự phân bố những khác biệt di truyền trong các quần thể và những thay đổi của sự phân bố đó theo thời gian[79]. Thay đổi về tần số một allele trong quần thể có thể là do ảnh hưởng của chọn lọc tự nhiên, khi tỷ lệ những cá thể mang một allele nào đấy sống sót và sinh sản được cao hơn khiến allele này xuất hiện nhiều hơn trong quần thể qua thời gian[80]. Sự biến động di truyền (genetic drift) cũng có thể diễn ra, khi những sự kiện bất chợt làm biến đổi ngẫu nhiên tần số allele[81].
Trải qua nhiều thế hệ, bộ gen của những sinh vật hoàn toàn có thể đổi khác, dẫn đến hiện tượng kỳ lạ tiến hóa. Đột biến và tinh lọc những đột biến có lợi giúp những loài tiến hóa và sống sót tốt hơn trong thiên nhiên và môi trường của chúng, quy trình này gọi là thích nghi [ 82 ]. Những loài mới được tạo thành trải qua quy trình hình thành loài, quy trình thường có nguyên do từ cách biệt địa lý dẫn đến những quần thể khác nhau trở nên cách ly về di truyền [ 83 ]. Việc ứng dụng những nguyên tắc di truyền vào nghiên cứu sinh học quần thể và tiến hóa được xem là thuyết tiến hóa tổng hợp tân tiến .Khi những trình tự được cách ly và đổi khác trong quy trình tiến hóa, những độc lạ giữa những trình tự hoàn toàn có thể được dùng như một đồng hồ đeo tay phân tử để tính khoảng cách tiến hóa giữa chúng [ 84 ]. Những so sánh di truyền nhìn chung được xem như phương pháp đúng đắn nhất để diễn đạt mối liên hệ giữa những loài – một tân tiến so với việc so sánh những đặc tính kiểu hình vốn dễ nhầm lẫn trước đây. Khoảng cách tiến hóa giữa những loài hoàn toàn có thể được phối hợp tạo thành cây tiến hóa – những cây này miêu tả nguồn gốc chung và sự phân hướng của những loài qua thời hạn, dù chúng không biểu lộ được sự chuyển giao vật tư di truyền giữa những loài không tương quan với nhau ( được biết đến là sự chuyển gen ngang và hầu hết phổ cập ở vi trùng ) .
Nghiên cứu và công nghệ tiên tiến[sửa|sửa mã nguồn]
Sinh vật mẫu[sửa|sửa mã nguồn]
 Drosophila melanogaster), một sinh vật mẫu phổ biến trong nghiên cứu di truyền học.Ruồi giấm thường ( ), một sinh vật mẫu phổ cập trong điều tra và nghiên cứu di truyền học .Dù những nhà di truyền học bắt đầu nghiên cứu và điều tra tính di truyền ở phong phú những loài sinh vật, nhưng sau đó họ mở màn điều tra và nghiên cứu tập trung chuyên sâu tính di truyền ở nhóm những sinh vật đặc biệt quan trọng. Thực tế, những điều tra và nghiên cứu quan trọng ở một sinh vật nhất định sẽ khuyến khích những nhà nghiên cứu sau đó lựa chọn sinh vật đó để tăng trưởng nghiên cứu và điều tra xa hơn, và như thế sau cuối chỉ có 1 số ít ít sinh vật mẫu đã trở thành cơ sở cho hầu hết những điều tra và nghiên cứu về di truyền [ 85 ]. Các đề tài điều tra và nghiên cứu di truyền phổ cập trên những sinh vật mẫu gồm có nghiên cứu và điều tra về điều hòa gen, mối tương quan giữa gen với sự tăng trưởng hình thái và ung thư .
Drosophila melanogaster), một sinh vật mẫu phổ biến trong nghiên cứu di truyền học.Ruồi giấm thường ( ), một sinh vật mẫu phổ cập trong điều tra và nghiên cứu di truyền học .Dù những nhà di truyền học bắt đầu nghiên cứu và điều tra tính di truyền ở phong phú những loài sinh vật, nhưng sau đó họ mở màn điều tra và nghiên cứu tập trung chuyên sâu tính di truyền ở nhóm những sinh vật đặc biệt quan trọng. Thực tế, những điều tra và nghiên cứu quan trọng ở một sinh vật nhất định sẽ khuyến khích những nhà nghiên cứu sau đó lựa chọn sinh vật đó để tăng trưởng nghiên cứu và điều tra xa hơn, và như thế sau cuối chỉ có 1 số ít ít sinh vật mẫu đã trở thành cơ sở cho hầu hết những điều tra và nghiên cứu về di truyền [ 85 ]. Các đề tài điều tra và nghiên cứu di truyền phổ cập trên những sinh vật mẫu gồm có nghiên cứu và điều tra về điều hòa gen, mối tương quan giữa gen với sự tăng trưởng hình thái và ung thư .
Các sinh vật được lựa chọn, một phần bởi tính thuận tiện: có vòng đời ngắn và dễ dàng thao tác di truyền. Những sinh vật mẫu được sử dụng rộng rãi bao gồm: vi khuẩn đường ruột Escherichia coli, cải Arabidopsis thaliana, men bánh mỳ Saccharomyces cerevisiae, giun tròn Caenorhabditis elegans, ruồi giấm (Drosophila melanogaster) và chuột nhà (Mus musculus).
Di truyền y học[sửa|sửa mã nguồn]
Di truyền y học tìm hiểu xem biến đổi di truyền liên hệ tới sức khỏe và bệnh tật của con người như thế nào[86]. Khi tìm kiếm một gen chưa biết mà có thể liên quan tới một căn bệnh, các nhà nghiên cứu thường sử dụng liên kết gen và sơ đồ phả hệ di truyền để tìm ra vị trí của nó trong bộ gen. Ở cấp độ quần thể, các nhà nghiên cứu lợi dụng sự ngẫu nhiên hóa Mendel (Mendelian randomization) để tìm ra những vị trí trong bộ gen mà liên đới với căn bệnh, một kỹ thuật đặc biệt hữu ích với những tính trạng đa gen không được xác định rõ ràng bởi một gen đơn lẻ[87]. Khi một gen tương ứng được tìm ra, những nghiên cứu xa hơn sẽ tiếp tục thực hiện với cùng gen đó (được gọi là các gen trực giao) trên những sinh vật mẫu. Bên cạnh nghiên cứu các bệnh di truyền, việc tăng tính hữu hiệu của các kỹ thuật kiểu gen đã đưa đến lĩnh vực di truyền học dược lý—nghiên cứu làm sao kiểu gen có thể tác động lên các phản ứng thuốc.[88]
Dù không di truyền được, ung thư vẫn được công nhận là một căn bệnh di truyền[89]. Quá trình phát triển ung thư của một cơ thể là sự kết hợp của nhiều sự kiện. Các đột biến thỉnh thoảng diễn ra trong các tế bào của cơ thể khi chúng phân chia. Trong khi những đột biến này sẽ không di truyền được sang thế hệ sau, chúng lại có thể tác động lên hoạt động của các tế bào, có khi khiến tế bào phát triển và phân chia nhanh hơn. Có những cơ chế sinh học cố gắng ngăn chặn quá trình này; những tín hiệu được chuyển đi tới những tế bào phân chia không thích hợp và khởi động quá trình apoptosis (tế bào chết theo chương trình). Tuy vậy, đôi lúc những đột biến thêm tiếp tục diễn ra làm tế bào không nhận được các tín hiệu. Một quá trình chọn lọc tự nhiên xảy ra bên trong cơ thể, và rốt cuộc, đột biến tích lũy trong các tế bào làm đẩy mạnh sự phát triển của chúng, tạo ra khối u ung thư, tiếp tục phát triển và xâm chiếm các mô khác nhau trong cơ thể sinh vật.
Kỹ thuật di truyền[sửa|sửa mã nguồn]
DNA hoàn toàn có thể được thao tác trong phòng thí nghiệm. Các enzym cắt số lượng giới hạn là loại enzym thường được sử dụng để cắt DNA thành những chuỗi riêng không liên quan gì đến nhau, tạo ra những đoạn DNA hoàn toàn có thể định trước được [ 90 ]. Việc sử dụng những enzym gắn cho phép những đoạn này nối lại với nhau, và nối những đoạn DNA từ những nguồn khác nhau ; nhờ thế những nhà nghiên cứu hoàn toàn có thể tạo ra DNA tái tổng hợp. Thường gắn liền với những sinh vật biến đổi gen, DNA tái tổng hợp thường thì được tạo nên từ những plasmid – những đoạn DNA vòng ngắn tiềm ẩn một vài gen. Bằng cách chèn plasmid vào vi trùng và nuôi những vi trùng này trên đĩa thạch agar ( để phân lập những dòng tế bào vi trùng ), những nhà nghiên cứu hoàn toàn có thể khuếch đại vô tính những đoạn DNA đã chèn ( quy trình được biết đến là tách dòng phân tử ) .DNA cũng hoàn toàn có thể được khuếch đại nhờ sử dụng một kỹ thuật gọi là phản ứng chuỗi trùng hợp ( PCR ). [ 91 ] Sử dụng những chuỗi DNA ngắn đặc hiệu, PCR hoàn toàn có thể phân lập và khuếch đại theo hàm mũ một vùng DNA đã xác lập. Bởi năng lực phóng đại kể cả những đoạn cực nhỏ của DNA, PCR liên tục được sử dụng để phát hiện sự xuất hiện của những trình tự DNA đơn cử .
Xác định trình tự DNA và hệ gen học[sửa|sửa mã nguồn]
Là một trong những kỹ thuật chủ yếu được phát triển để nghiên cứu di truyền học, “xác định trình tự DNA” (DNA sequencing) cho phép các nhà nghiên cứu xác định trình tự nucleotide trên một đoạn DNA. Phát triển năm 1977 bởi Frederick Sanger và các cộng sự, phương pháp xác định trình tự gián đoạn chuỗi hiện nay là phương pháp được sử dụng thường lệ[92]. Với kỹ thuật này, các nhà khoa học có thể nghiên cứu được những trình tự phân tử liên quan tới nhiều bệnh di truyền ở người.
Khi xác định trình tự đã trở nên đỡ tốn kém hơn, cùng với sự trợ giúp của các công cụ tính toán, những nhà nghiên cứu đã xác định được bộ gen của nhiều sinh vật bằng cách liên kết trình tự của nhiều đoạn khác nhau (quá trình này gọi là “lắp ráp bộ gen” – genome assembly)[93]. Những kỹ thuật trên được sử dụng để xác định bộ gen người, đã được hoàn thiện trong Dự án bản đồ gen người vào năm 2003[39]. Những kỹ thuật xác định trình tự cao năng (high-throughput) mới đột ngột làm giảm chi phí xác định trình tự DNA, đem tới hy vọng mới cho nhiều nhà nghiên cứu rằng có thể thực hiện được việc này với giá thành chỉ còn 1000 đô la Mỹ[94].
Thành tựu giải trình tự DNA ngày càng nhiều, kết hợp với các nhu cầu khác của y học, dân tộc học v.v đã thúc đẩy sự hình thành nên hệ gen học (genomics) – một khoa học liên ngành nghiên cứu về tất cả các gen của bộ gen trong cơ thể, sử dụng các công cụ tính toán để tìm kiếm và phân tích các mô hình trong bộ gen đầy đủ của sinh vật. Hệ gen học có ứng dụng và liên quan nhiều đến tin sinh học, là một bộ môn sử dụng máy tính cùng những thuật toán khác để tập hợp các dữ liệu sinh học cũng như mô phỏng các quá trình sinh học.
Một vài yếu tố xã hội tương quan[sửa|sửa mã nguồn]
Có nhiều yếu tố về di truyền học tương quan đến xã hội, đang được bàn cãi .
Sự di truyền trí mưu trí[sửa|sửa mã nguồn]
Trí mưu trí loài người có mang tính di truyền hay không là một yếu tố được tranh cãi và điều tra và nghiên cứu kể từ khi di truyền học sinh ra cho đến nay và hoàn toàn có thể còn liên tục lê dài. Đa số những học giả chấp thuận đồng ý rằng tính di truyền có tác động ảnh hưởng nhất định đến sự mưu trí, tuy nhiên ở mức độ nào thì vẫn còn nhiều tranh luận. Sự phức tạp của yếu tố tăng lên khi một số ít fan hâm mộ đưa thêm yếu tố chủng tộc vào. Một số học giả nhìn nhận di truyền trí mưu trí qua chỉ số mưu trí ( IQ ), 1 số ít khác cho rằng yếu tố phức tạp hơn nhiều, không hề chỉ đơn thuần địa thế căn cứ vào IQ. [ 95 ] Thêm vào đó một số ít người lại cho rằng trí mưu trí được di truyền theo mẹ .
Ưu sinh học[sửa|sửa mã nguồn]
Thuật ngữ ưu sinh học (eugenics) được nêu ra lần đầu vào năm 1893, với mục tiêu phát triển lĩnh vực “cải thiện giống người”. Tuy vậy, sau khi bị Đức Quốc xã lợi dụng vào những năm 1920-1930, ưu sinh học đã không được nhắc tới trong thời gian dài. Người ta phân ra hai hình thức của ưu sinh học:
- Ưu sinh học âm: mục tiêu là giảm tần số các gen xấu. Chẳng hạn như có một số nước cấm những người mắc bệnh di truyền không được sinh con.
- Ưu sinh học dương: mục tiêu là tăng tần số các gen tốt. Chẳng hạn như ở Mỹ hằng năm có khoảng 5.000-10.000 trẻ em được sinh ra từ tinh trùng của những người cha được chọn lọc.
Hiện nay, một số ít người cho rằng hoàn toàn có thể dùng liệu pháp gen để tái tạo loài người, nhưng chỉ nên tác động ảnh hưởng ở tế bào soma. Việc tái tạo con người hay tạo ra những con người siêu việt vấp phải một số ít yếu tố đạo đức và nhân chủng học. [ 96 ]
Đạo đức sinh học[sửa|sửa mã nguồn]
Việc tăng trưởng của ngành sinh học nói chung và di truyền học nói riêng đã tạo ra không ít yếu tố về đạo đức sinh học. Một số nhà khoa học đã đề xuất thực thi trưng cầu dân ý để cấm việc nghiên cứu và điều tra về di truyền học. Một số khác thì đề xuất cần có những luật lệ rõ ràng để bảo vệ bộ gen người, tương thích với những quy chuẩn đạo đức của con người. Nhiều yếu tố tâm ý xã hội phát sinh khi biết rõ bộ gen một người nào đó : nếu họ là những người thông thường nhưng có mang gen bệnh thì yếu tố hôn nhân gia đình, sinh đẻ hay xin việc làm của họ sẽ như thế nào .Sau khi thí nghiệm về nhân bản người được thực thi tại Mỹ năm 1993, 1 số ít nước thuộc Cộng đồng châu Âu đã đưa ra những luật lệ cấm những thí nghiệm dạng này. Một số nhà bảo vệ môi sinh kịch liệt phản đối việc nhân bản người và sinh vật bằng những kỹ thuật di truyền. [ 97 ]UNESCO đã lập ra Ủy ban quốc tế về Đạo đức sinh học nhằm mục đích thu nhập những quan điểm kiến thiết xây dựng nên những luật lệ về đạo đức sinh học tương quan đến bộ gen người .
Sinh vật đổi khác di truyền[sửa|sửa mã nguồn]
Kỹ thuật di truyền tạo ra những sinh vật biến hóa về mặt di truyền ( GMO ) đã khiến nhiều người quan ngại : liệu những sinh vật này có ép chế, ảnh hưởng tác động xấu tới những dạng sinh vật tự nhiên khác hay tạo ra những dạng bệnh mới do tái tổng hợp với những dạng tự nhiên ? Liệu những gen của những sinh vật đổi khác di truyền có gây nguy hại cho khung hình con người hay không ? Liệu những thực vật kháng chất diệt cỏ ví dụ điển hình có năng lực chuyển gen cho cỏ dại hay không. v.v. Trước những quan ngại này, nhiều nước đã kiến thiết xây dựng luật lệ ngặt nghèo để trấn áp những sinh vật GMO. [ 98 ]
Khoa học hình sự[sửa|sửa mã nguồn]
Do mọi mô trong khung hình đều chứa cấu trúc DNA nguyên vẹn của một thành viên nên khoa học pháp y hoàn toàn có thể dựa vào những mẫu sinh học tìm thấy ở hiện trường để kiến thiết xây dựng được hồ sơ di truyền học của khung hình đó, từ đó giúp xác lập thủ phạm hay vô hiệu nghi can vô tội trong một vụ án. Ngoài ra, những phép nghiên cứu và phân tích di truyền cũng được cho phép chứng minh và khẳng định hay loại trừ một nghi vấn về quan hệ huyết thống nào đó, ví dụ điển hình như trong trường hợp xác lập cha mẹ của một đứa trẻ bị thất lạc. [ 99 ]
Liên kết ngoài[sửa|sửa mã nguồn]
Source: https://dvn.com.vn
Category: Hỏi Đáp